

今天要和大家分享的内容是免疫细胞家族中的B细胞的相关研究思路~
一、相关概念
B细胞(B cells),也称B淋巴细胞(B lymphocytes),是淋巴细胞亚家族白细胞中一员。B细胞主要功能是在体液免疫中分泌抗体,可以在其细胞膜表面表达B细胞受体,结合抗原(进入体内并使您生病的有害病原体,例如病毒、细菌和寄生虫),后诱导抗体(针对侵袭性病原体特异性免疫球蛋白Ig)的产生,以抵御这些病原体的入侵。此外,B细胞也可以呈递抗原和分泌细胞因子(包括CCL22/17、IL-2/4/6、INF-γ、TNF-α、GM-CSF、TGF-1β、IL-10/35)。
二、B细胞在哺乳动物中的发育路径
在哺乳动物中,B细胞在骨髓中发育,随后在次级淋巴组织中功能成熟,并最终在血液中循环。
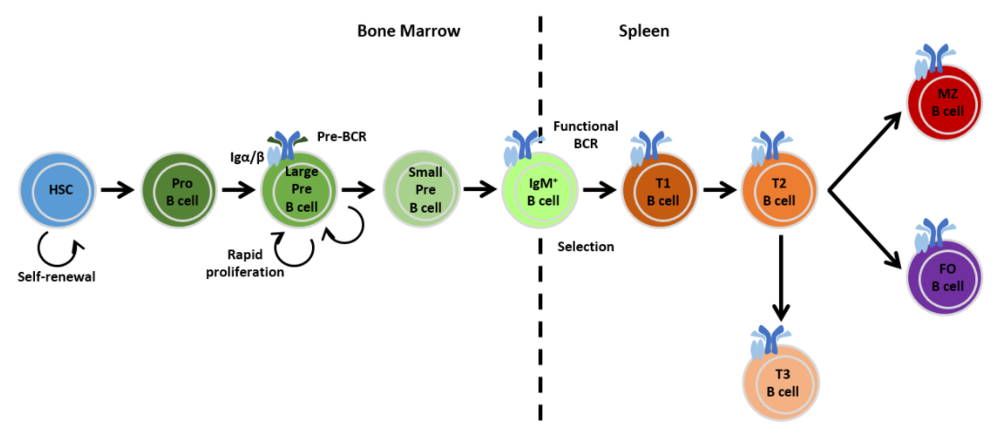
01成熟与未成熟B细胞
B细胞源于始于骨髓的造血干细胞系列。在骨髓中,造血干细胞系列首先分化为多能祖细胞(HSC),再分化为祖B细胞(pro B cells),表达lgα/β异源二聚体。BCR是存在于B细胞膜表面的免疫球蛋白(Ig),由两条相同的重链(HC)和两条相同的轻链(LC)组成。在B细胞发育过程中,免疫球蛋白的这两条链会进行序贯基因重排,期间先在细胞表面生成pre-BCR。随后,HC和LC基因成功重排的早期B细胞(pre B cells)成为表达膜IgM功能性BCR的未成熟B细胞。未成熟B细胞和成熟的B细胞之间的区别就在于,成熟B细胞能够响应抗原刺激并产生抗体,而未成熟B细胞则不能。
02两轮筛选
未成熟的B细胞想要变成成熟的B细胞,需要在骨髓中进行两轮筛选——阳性选择和阴性选择。
阳性选择的结果与抗原反应无关,仅仅是针对B细胞受体(BCR)以及其前体pre-BCR的。如果这些受体不能很好地结合它们的配体,B细胞就会停止发育。
而阴性选择的目的,是检查B细胞与自身抗原的结合力,如果结合力过强,B细胞就会凋亡。由于未成熟B细胞可能会对自体抗原产生免疫发应,这轮选择就能够确保B细胞的免疫耐受的状态,使得其不会对自体抗原发生免疫反应。
经历了两轮选择的未成熟B细胞就可以进入下一步的发育,即从骨髓迁移到脾脏成为过渡型B细胞。
03过渡型B细胞
经历了两轮选择的未成熟B细胞就可以进入下一步的发育,即从骨髓迁移到脾脏成为过渡型B细胞。
过渡型B细胞分两个阶段:T1和T2。B细胞迁入至脾脏后为T1型B细胞,TI细胞会在脾脏内完成转换,形成T2型B细胞。
等到T1转换成T2以后,根据受体接收到的信号的不同,T2型B细胞既可以分化为滤泡(follicular)B细胞,也可以分化为边缘区(marginal zone)B细胞。一旦分化完成,它们就被视为成熟(mature)B细胞,或者说初级(naive)B细胞。成熟了的B细胞会主要定居在淋巴结皮质浅层的淋巴小结和脾脏的红髓和白髓的淋巴小结内。
三、案例分析
接下来,我们将通过一篇高分文献(“LAPTM5通过调控WWP2-PTEN-AKT通路介导未成熟B细胞凋亡和B细胞耐受”),带大家了解作者的课题思路。

01研究背景
B细胞发育是免疫球蛋白(Ig)HC和LC基因重排的过程。由于重排的随机性,据估计,多达85%的新生成的未成熟B细胞可以识别自身抗原。
B细胞中枢耐受功能通过至少三种机制去除骨髓(BM)中的自身反应性未成熟B细胞:
① 克隆清除:诱导以高亲和力识别自身抗原的未成熟B细胞的凋亡”;
② 或者,这些未成熟的B细胞可能启动新一轮的轻链重排,这一过程被称为受体编辑,以改变BCR的特异性;
③ 由于BCR表达信号的下调,未成熟的B细胞对自身抗原的识别度较低,可能会在功能上无反应或无能。
为了防止这种自身反应性B细胞产生自身抗体,免疫系统已经开发出多种机制,包括B细胞中枢和外周耐受,以消除/灭活自身反应性B细胞。诱导识别自身抗原的未成熟B细胞的凋亡是B细胞中枢耐受去除这些未成熟B细胞的一个重要机制。然而,BCR信号如何触发未成熟B细胞的凋亡的机制鲜有人知。
溶酶体相关蛋白跨膜5(LAPTM5)是一种在淋巴细胞和髓细胞中表达的蛋白,它包含五个跨膜结构域、三个多聚脯氨酸酪氨酸(PY)基序和一个泛素相互作用(UIM)基序。已有研究发现LAPTM5负调节成熟B细胞的表面BCR水平,而LAPTM5缺乏导致B细胞活化和自身抗体产生增强,此外LAPTM5还介导早期B谱系细胞中pre-BCR的下调。但LAPTM5是否能调节BCR信号从而引发未成熟B细胞的凋亡是未被确认的。
Fig 1:与56R小鼠相比,Laptm5−/−56R小鼠骨髓(BM)中未成熟B细胞群增加
首先,作者为了探究Laptm5对未成熟B细胞群数量的影响,将56R HC敲入小鼠体内。56R HC与大多数内源性轻链配对时,以高亲和力结合DNA。只有少数轻链,称为编辑器,能够抑制/减少DNA结合,并允许未成熟B细胞的存活和成熟。结果发现Vκ21D(编辑器轻链)消除了DNA结合,而Vκ38C(编辑器轻链)允许低亲和力的DNA结合。
因此,56R敲入小鼠适用于研究同一体内系统中高亲和力和低亲和力自身反应性B细胞的生成和命运,于是作者比较了56R小鼠和Laptm5−/−56R小鼠骨髓细胞中的未成熟B细胞群数量。
流式细胞术分析结果显示,在56R小鼠中,再循环B细胞的百分比和数量降低;但在B220+CD43−细胞中,56R和Laptm5−/−56R小鼠的pre-B细胞的比例增加。
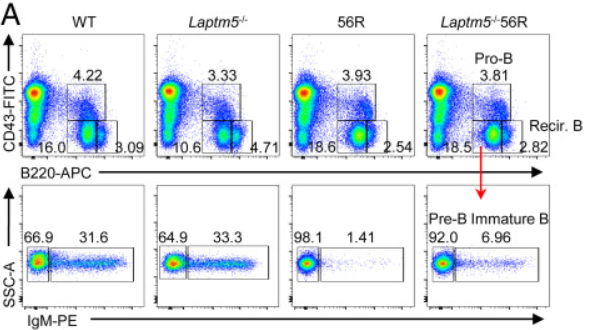
且与WT小鼠相比,56R小鼠的未成熟B细胞比例和数量显著减少,因为大多数携带56R HC的未成熟B细胞是自反应性的,因此它们被消除了。值得注意的是,与56R小鼠相比,Laptm5−/−56R小鼠的未成熟B种群显著增加。
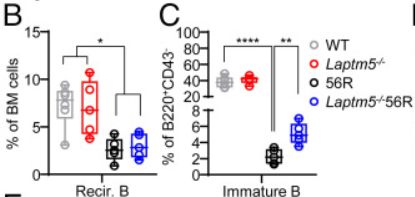
这也就说明了LAPTM5缺陷允许一些本会因凋亡而被删除的自身反应性未成熟B细胞存活下来。
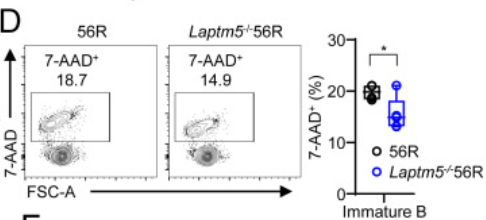
并且通过再一次的对比,我们也可以看出,Laptm5−/−56R小鼠,未成熟的B细胞中7-AAD+死细胞的比例明显低于56R小鼠中该类死细胞的数量。
为了进一步揭示LAPTM5在BM中B细胞发育中的作用,作者对56R和Laptm5−/−56R小鼠的BM B220+细胞进行了单细胞RNA测序(scRNA-Seq),并通过无监督分层聚类和均匀流形近似和投影(UMAP)可视化,将这些细胞划分为11个独特的簇群。
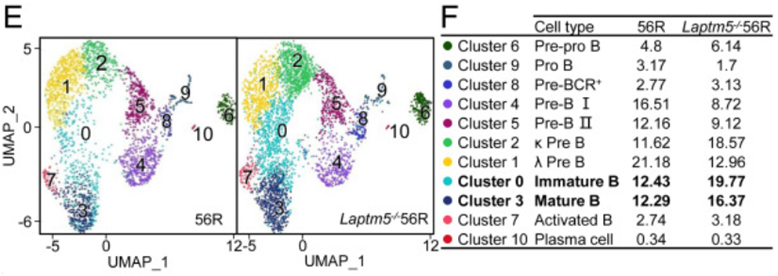
测序结果与荧光激活细胞分选数据一致,即与56R小鼠相比,Laptm5−/−56R小鼠含有更高比例的未成熟B细胞和成熟B细胞。
以上这一部分的实验,证明了Laptm5缺陷会使小鼠骨髓(BM)中未成熟B细胞群增加。
Fig 2:与56R小鼠相比,Laptm5−/−56R小鼠的自身反应性B细胞增加,自身抗体水平升高
接着,作者通过比较了56R小鼠和Laptm5−/−56R小鼠骨髓(BM)中的成熟B细胞分布,结果如图显示,通过这张图我们可以看出,与56R小鼠相比,来自Laptm5−/−56R小鼠成熟B细胞富集于激活的自身免疫性疾病和移植排斥途径,这表明Laptm5−/−56R小鼠可能更容易获得自身免疫。
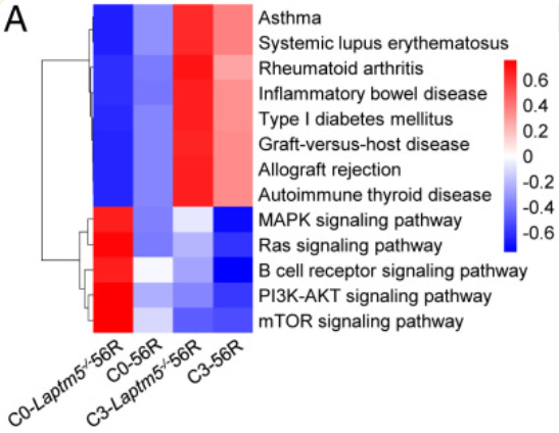
于是,作者进一步比较了表达56R HC的未成熟和成熟B细胞的LC谱。
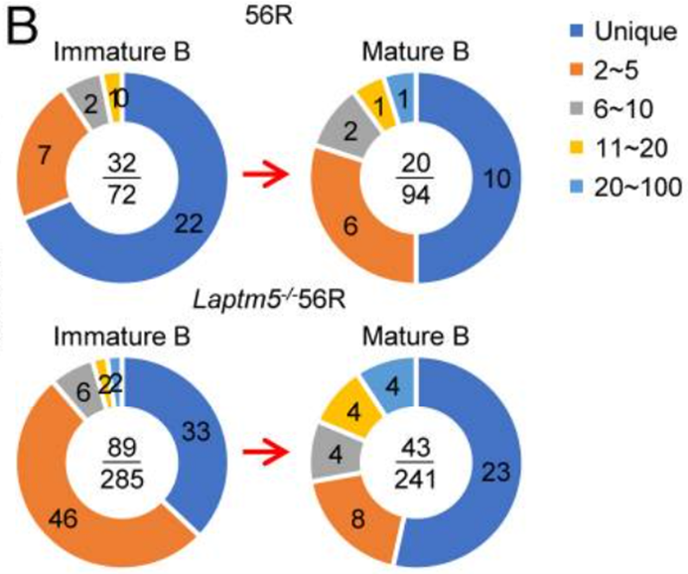
在左上谱中,用于实验的56R小鼠的未成熟B细胞中72个LC序列中,发现了32个不同的序列。
相比之下,右下来自成熟B细胞的94个LC序列仅包含20个独特的序列,与56R小鼠的未成熟B细胞相比,成熟B细胞的LC多样性减少了52%。
在Laptm5−/−56R小鼠中,未成熟B细胞中的285个LC序列中有89个是独特的(左下),而成熟B细胞中241个LC序列中的43个是独特的(右下),相当于成熟B细胞中LC多样性降低了43%。
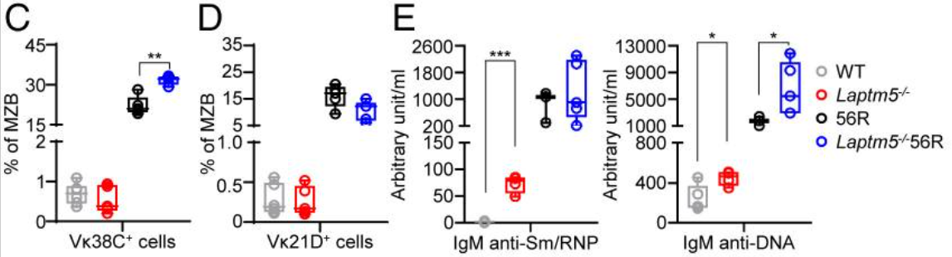
此外,作者还发现,与56R小鼠相比,Laptm5−/−56R小鼠含有更高比例的Vκ38C+ MZB细胞,同时Vκ21D+ MZB细胞比例减少,显示LAPTM5缺乏使更多的Vκ38C+自身反应性B细胞成为MZB细胞。而Laptm5−/−小鼠产生的抗smrnp和抗dna自身抗体水平均高于WT小鼠。并且,Laptm5−/−56R小鼠比56R小鼠产生更高水平的IgM抗DNA和IgM抗核抗体。
这些结果表明LAPTM5在限制自身反应性B细胞转变为MZB细胞和分泌自身反应性IgM抗体方面具有重要作用。
通过这一部分实验,作者得出结论:Laptm5缺失能够使小鼠自身反应性B细胞增加,自身抗体水平升高。
Fig 3:BCR交联部分通过上调LAPTM5诱导未成熟B细胞凋亡
接下来,作者利用未成熟B细胞系WEHI231模型,对Laptm5介导的未成熟B细胞凋亡的作用进行探究,实验结果如图所示。
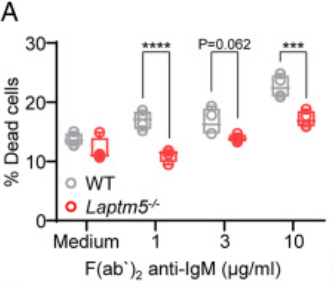
图A说明与WT未成熟B细胞相比,LAPTM5缺失的未成熟B细胞显示出更少的细胞死亡,这也表明了LAPTM5缺失使未成熟B细胞更能抵抗BCR刺激诱导的死亡。图B-C说明,通过使用α-IgM处理上调未成熟B细胞中LAPTM5蛋白的表达,将会触发细胞死亡。
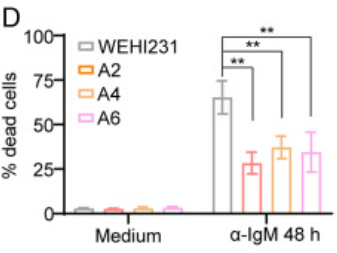
接着在与WT型WEHI231细胞相比后,作者发现:所有三个LAPTM5缺陷克隆的凋亡均显著减少,这意味着α-IgM诱导的细胞凋亡部分是由LAPTM5介导的。
而在图E显示的实验结果中,我们可以发现一个有趣的现象,在WEHI231细胞中,SYK、ERK和AKT的磷酸化在BCR结合后的24和48小时急剧减少;在α-IgM刺激的WEHI231细胞中,p27Kip1和BIM的表达均上调。与WEHI231细胞类似,LAPTM5缺陷的WEHI231细胞(A2)呈现下调的p-SYK水平。
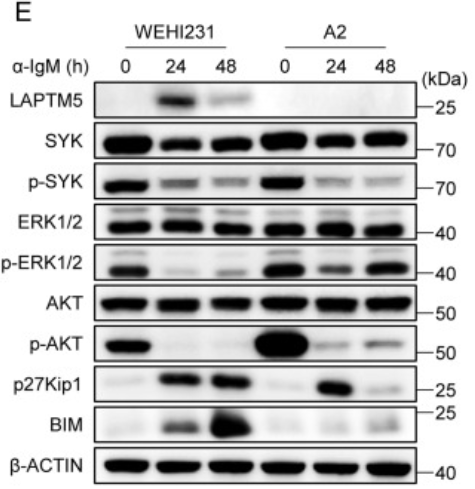
然而,A2细胞在α-IgM刺激前后的p-ERK和p-AKT水平高于WEHI231细胞,并且在α-IgM刺激后48小时仅适度上调p27Kip1和BIM蛋白表达。与WEHI231和A2细胞的研究结果一致,LAPTM5缺陷的BM未成熟B细胞与WT BM未成熟B细胞相比,p-SYK水平相似,但p-AKT水平更高。
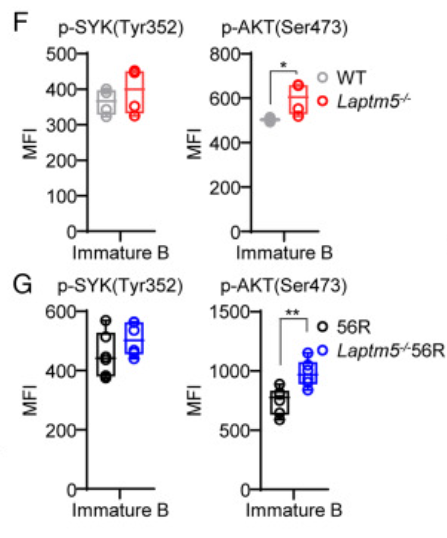
且与56R未成熟B细胞相比,Laptm5−/−56R未成熟B细胞显示出相似水平的p-SYK,但p-AKT水平更高。这足以表明LAPTM5负调节WEHI231和BM未成熟B细胞中的p-AKT水平。
随后,作者进行了进一步实验,实验结果如图H和I显示,图H表明LAPTM5诱导的细胞凋亡伴随着线粒体膜电位低的细胞百分比增加。而图I表明在LAPTM5表达后48小时降低p-SYK和p-ERK水平;同时,p-AKT水平降低,而p27Kip1和BIM水平均上调。信号分子磷酸化和p27Kip1上调的动力学略有延迟可能是由于逆转录病毒转导后LAPTM5表达所需的时间。此外,我们可以发现FOXO1在LAPTM5表达后24小时上调,而抗凋亡MCL-1的表达在48小时降低。BAX和BCL-xL的水平不受影响。
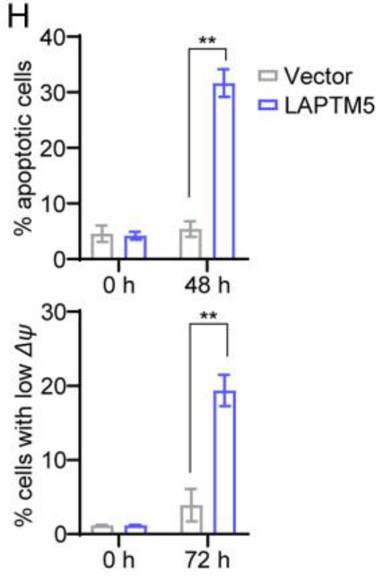
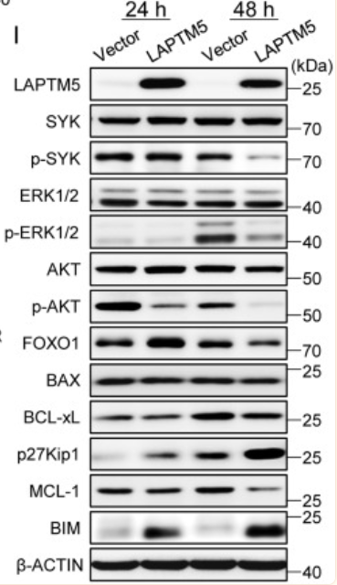
因此作者在这部分实验中得出结论:BCR交联部分通过上调LAPTM5诱导未成熟B细胞凋亡。
Fig 4:LAPTM5通过上调PTEN水平来抑制未成熟B细胞中AKT磷酸化
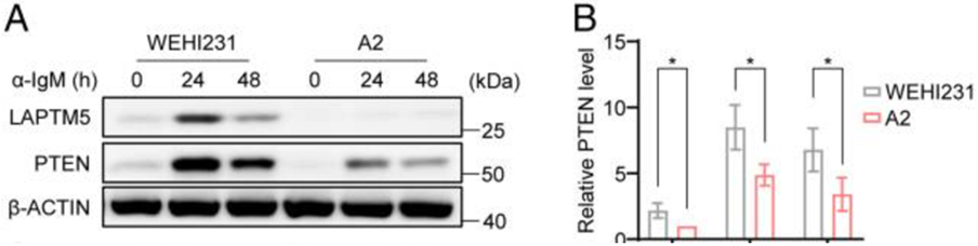
根据图A-B显示的结果,可以发现A2细胞的PTEN基础水平低于WEHI231细胞(0h)。α-IgM刺激上调了WEHI231细胞的LAPTM5蛋白表达,但A2细胞中的没有上调,在24h和48h时,A2细胞的PTEN水平也低于WEHI231细胞中PTEN水平。
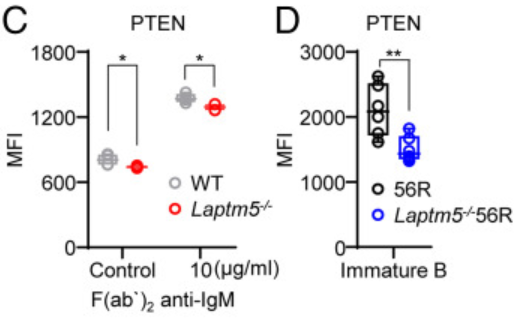
而图C-D的结果说明了与WT BM未成熟B细胞相比,Laptm5−/−BM未成熟B细胞在α-IgM刺激前后表达的PTEN水平较低。此外,Laptm5−/−56R未成熟B细胞的PTEN水平也低于56R未成熟B细胞的PTEN水平。
这些结果可以说明-IgM刺激可能通过LAPTM5上调PTEN蛋白水平。
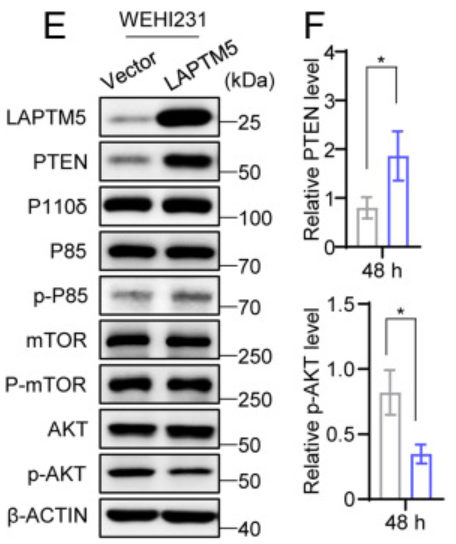
为了探索这种可能性,作者在WEHI231细胞中异位表达LAPTM5。
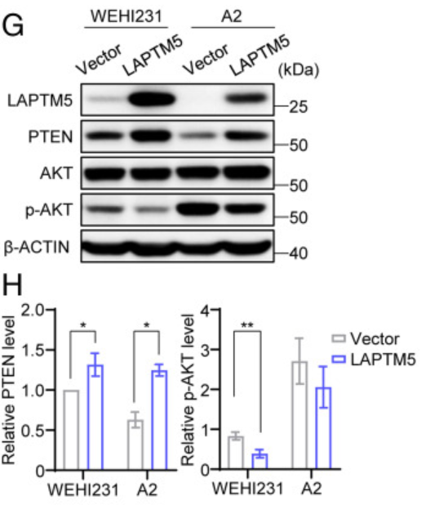
我们可以发现,LAPTM5的表达确实导致PTEN水平升高,而PI3K催化亚基P110δ和调节亚基P85以及mTOR和p-mTOR的水平则保持不变。于是,为了进一步验证LAPTM5与PTEN和p-AKT的关系,作者比较了WEHI231和A2细胞中的PTEN和p-AKT水平,得出以下结果。
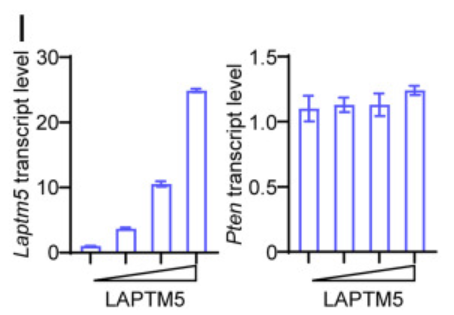
WEHI231细胞的PTEN水平更高,p-AKT水平低于A2细胞,表明内源性LAPTM5导致PTEN水平升高和p-AKT水平降低。此外,LAPTM5的异位表达导致WEHI231和A2细胞中PTEN水平升高和p-AKT水平降低。这些结果表明,LAPTM5上调PTEN水平,从而打破PI3K和PTEN活性之间的平衡,导致AKT磷酸化受抑制,且PTEN转录本的水平不受LAPTM5表达增加的影响。
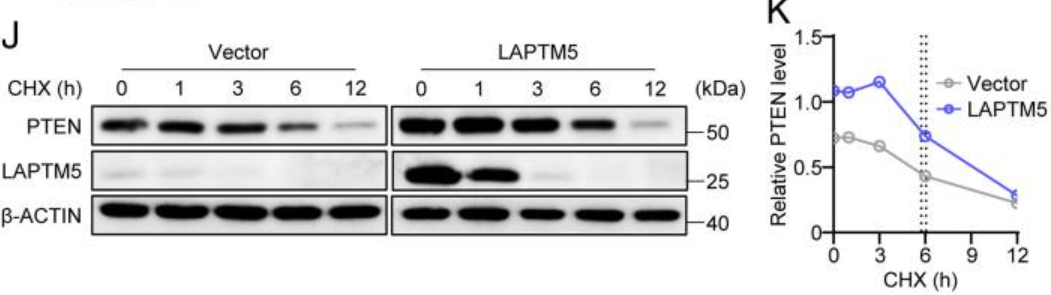
接着作者使用抑制新蛋白质合成的环己酰亚胺(CHX),并发现PTEN蛋白在表达LAPTM5的WEHI231细胞中的半衰期为6.06小时,而在对照细胞中的半衰期为5.74小时。
于是,作者得出结论:LAPTM5会通过上调PTEN水平来抑制未成熟B细胞中AKT磷酸化。
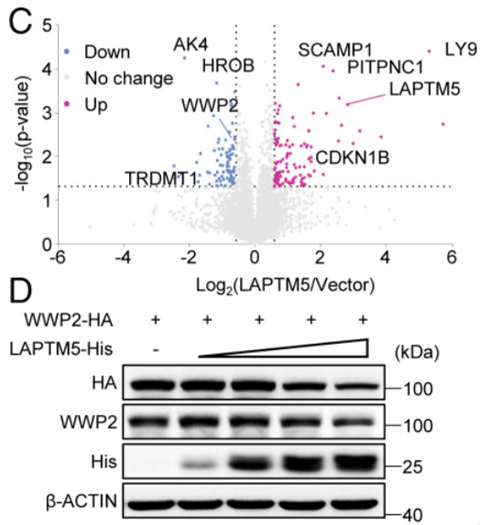
Fig 5:LAPTM5与WWP2相互作用并促进WWP2在溶酶体中的降解
为了阐明LAPTM5上调PTEN的机制,作者绘制了LAPTM5组和载体组的热图,分析显示189个蛋白质形成明显不同的簇,通过分层聚类可以确定,每组的3个重复显示出较低的重复间变异。
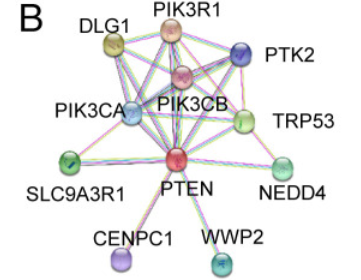
于是,作者采用相互作用基因检索工具(STRING)数据库,分析并显示出与PTEN相互作用的蛋白质包括三个PIK3家族成员(PIK3CA/B/R1)、DGL1、PTK2、TRP53、SLC9A3R1、CENPC1、NEDD4和WWP2。
通过进一步的实验验证,作者发现,LAPTM5能够降低WWP2表达,并且在HEK293细胞中,LAPTM5异位表达以剂量依赖的方式降低了WWP2蛋白水平。
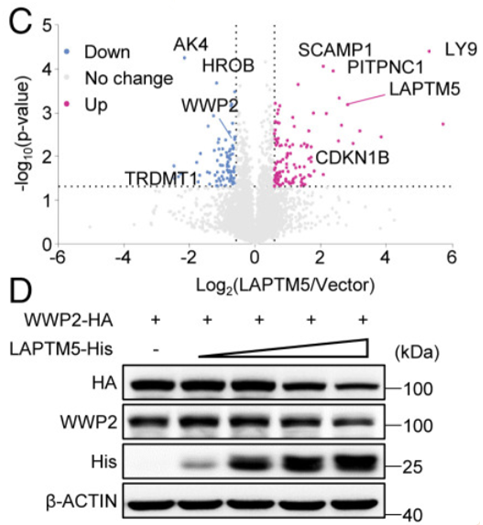
这一步的发现,揭示了LAPTM5与WWP2相互作用并促进WWP2在溶酶体中的降解。
Fig 6:LAPTM5通过PY基序与WWP2相互作用并促进其溶酶体降解,导致PTEN水平升高
为了确定LATPM5与WWP2相互作用的机制,及其对PTEN水平的影响,作者进行了进一步的实验。
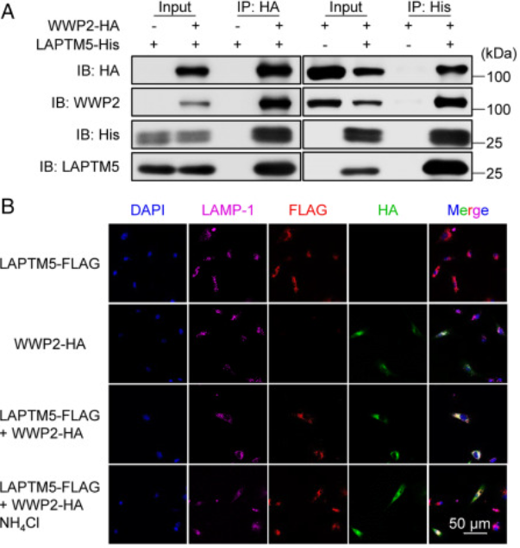
图A显示LAPTM5可与WWP2共沉淀,反之WWP2也能与LAPTM5共沉淀。于是作者通过共聚焦免疫荧光染色进一步分析了它们的共定位,发现LAPTM5和WWP2与溶酶体标志物LAMP1共定位。
而图C-D说明LAPTM5的异位表达会降低HEK293细胞中WWP2蛋白的水平,这种影响可以通过添加溶酶体活性抑制剂NH4Cl来消除,但蛋白酶体抑制剂MG132不能消除这种影响。
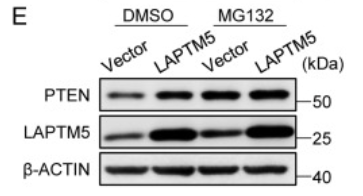
在MG132缺失的情况下,异位LAPTM5表达会增加PTEN蛋白水平。而在MG132存在时,载体转导细胞中的PTEN水平升高,这一系列的对比表明PTEN正常经历蛋白酶体降解。在之后,作者发现发现WWP2介导的PTEN泛素化被LAPTM5降低了。
于是作者假设LAPTM5也可能通过其PY基序与WWP2相互作用,并调节WWP2介导的PTEN泛素化。为了验证这一假设,作者进行了免疫沉淀和免疫印迹分析,结果证明,PTEN泛素化被WT LAPTM5消除,而三个PY基序全部突变(mPYs)的LAPTM5则没有。同样,mPYs未能提高PTEN蛋白水平。
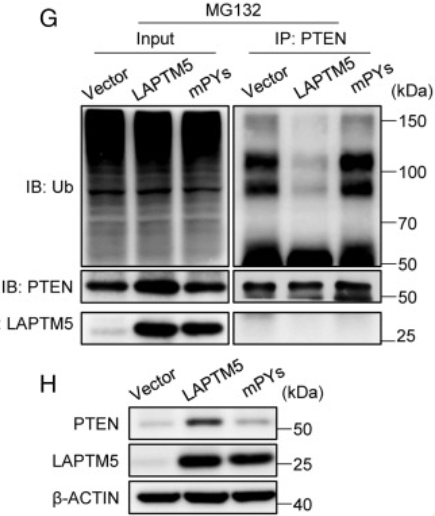
这些结果共同表明LAPTM5会通过PY基序与WWP2相互作用并促进其溶酶体降解,从而抑制WWP2介导的泛素化和PTEN的蛋白酶体降解,使PTEN的水平升高。
Fig 7:LAPTM5介导未成熟的B细胞凋亡并维持B细胞耐受性
作者通过结合本文结果及往研究进行推测和总结,绘制了这张图表,主要表明了LAPTM5的介导会让未成熟的B细胞凋亡并维持B细胞耐受性,这也证实了作者一开始的研究猜想。
02研究思路
Fig 1:使用56R HC敲入小鼠来阐明LAPTM5在消除和选择高和低亲和性自身反应性B细胞中的作用;
Fig 2:鉴定LAPTM5参与消除BM中自反应性未成熟B细胞,并限制低亲和性性自反应性B细胞成为MZB细胞的能力;
Fig 3:利用未成熟B细胞系WEHI231模型,明确LAPTM5介导的未成熟B细胞细胞凋亡的作用;
Fig 4-6:探讨LAPTM5介导的细胞凋亡的分子机制
Fig 7:结合本文结果及往研究进行推测和总结
这篇文章向我们揭示了一种消除自身反应性发育的B细胞以防止自身抗体产生的重要的机制。BCR刺激上调LAPTM5的表达,进而通过两种途径触发未成熟B细胞的凋亡,并致使SYK和ERK的磷酸化降低,进而可能导致PI3K与AKT磷酸化受抑制。
同时,LAPTM5靶向E3泛素连接酶WWP2介导溶酶体降解,导致其底物PTEN的积累。PTEN水平的升高抑制AKT磷酸化,导致FOXO1表达增加和细胞周期抑制剂p27Kip1和促凋亡分子BIM的上调。在体内,LAPTM5参与消除自身反应性B细胞,其缺乏会加剧自身抗体的产生。该结果显示LAPTM5-WWP2-PTEN级联反应有助于未成熟的B细胞凋亡和B细胞耐受。
四、研究思路
那么,我们应该怎样设计B细胞相关课的课题呢?
一般而言,B细胞研究一般包括:
一、确定B细胞促进自身免疫性疾病中的分子机制;
二、调查B细胞-T细胞相互作用与肿瘤进展的关系;
三、解析B细胞在免疫代谢中的作用;
四、借助基因编辑改造B细胞,使其持久表达的治疗性蛋白质。
 文章推荐
文章推荐

业务咨询
业务咨询专线:133 7682 0615
Email:lxyjy@wie-biotech.com


